Revistas
Revistas Archivo
Archivo









































Este artículo es parte de la edición de octubre, 2019
El poder sobrenatural de una microbiota intestinal saludable

El intestino de las aves es un ecosistema complejo y dinámico formado por microorganismos, bacterias, protozoos, hongos, levaduras, bacteriófagos y otros virus en el lumen y la mucosa.
Miles de especies bacterianas, hasta 1010 células bacterianas por gramo de digesta, habitan el intestino de las aves adultas. Este ecosistema es conocido como microbiota
Este complejo ecosistema crea una relación simbiótica con el huésped. Las interacciones entre la microbiota y la mucosa son esenciales para mejorar y proteger las funciones digestivas vitales. En individuos sanos, la microbiota está involucrada en la digestión de nutrientes, pero también en la prevención de la infección por microorganismos dañinos.
A. La instauración de la microbiota intestinal
Al nacer, el tracto digestivo del pollito joven es estéril.
En un sistema de producción comercial el ambiente de la incubadora es muy limpio y no contiene la microbiota digestiva de la gallina. Por tanto, la colonización microbiana depende de las fuentes ambientales para alcanzar su densidad total en un par de días -Apajalahti y col., 2004-.
La composición de esta flora fluctúa a lo largo de la vida de las aves según la nutrición, el ambiente, la estirpe, el sexo y los factores potenciales de estrés. Además, la población bacteriana es diferente en los diferentes tramos del intestino – Figura 1.
Figura 1.Composición de microbiota a lo largo del tracto digestivo del pollo
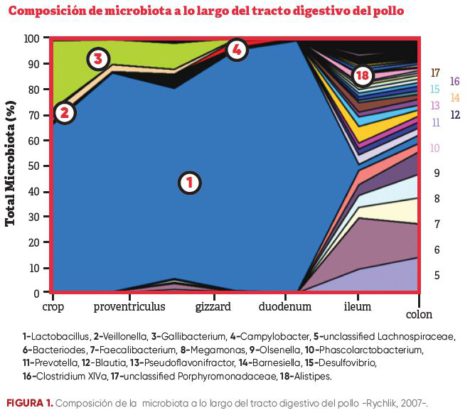
La suplementación de probióticos se han utilizado para modular la composición de la microbiota intestinal al competir con éxito con patógenos a través de un proceso de exclusión competitiva -Awaad y col., 2003-.
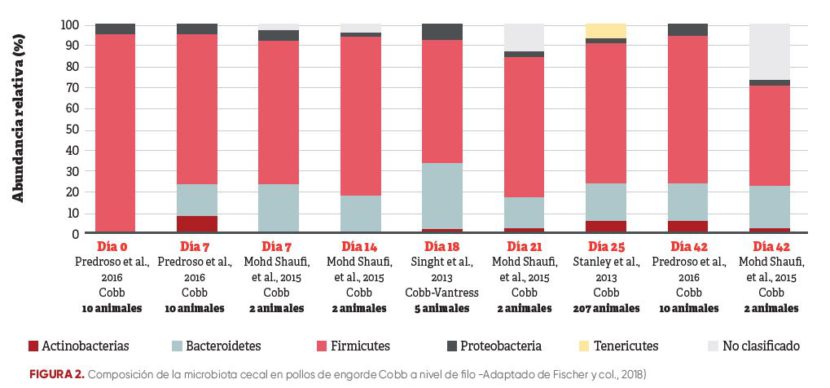
FIGURA 2. Composición de la microbiota cecal en pollos de engorde Cobb a nivel de filo -Adaptado de Fischer y col., 2018)
B. La acción microbiana a favor de la digestión de nutrientes
Las aves y su microbiota intestinal se benefician del intercambio de nutrientes huésped-microorganismo en una relación simbiótica.
En las aves sanas, la mayoría de los nutrientes digeribles se absorben y están disponibles para el metabolismo del huésped en el intestino delgado, donde la densidad (figura 3) bacteriana es menor.
En el ciego se da una fermentación bacteriana de carbohidratos más complejos, que no han sido digeridos enzimáticamente en la parte superior.
FIGURA 3. Interacciones entre la microbiota intestinal de las aves y el huésped para la disponibilidad de nutrientes
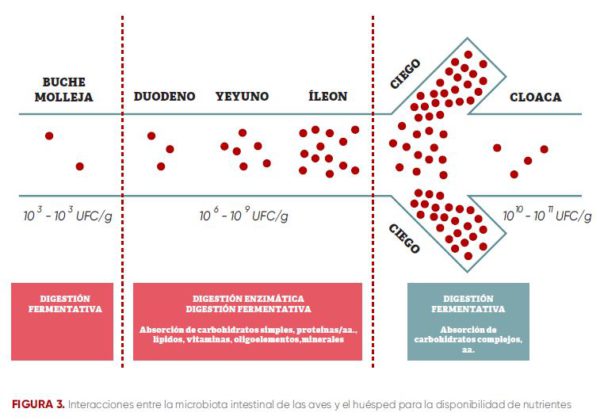
En condiciones normales, la microbiota intestinal contribuye a la disponibilidad de nutrientes de tres maneras distintas.
Las bacterias producen ácidos grasos de cadena corta -AGCC- y amoníaco, fuentes de energía y aminoácidos para el metabolismo del huésped.
Este proceso de fermentación es la primera acción directa de la microbiota sobre la absorción de nutrientes. El segundo efecto es la capacidad de afectar positivamente la digestibilidad del alimento al mejorar la actividad enzimática.
Al comparar la actividad de la enzima fosfatasa alcalina en pollos libres de gérmenes frente a pollos criados convencionalmente, observamos que los animales convencionales muestran una mayor actividad enzimática.
También se ha documentado que la levadura viva Saccharomyces cerevisiae boulardii tiene la capacidad de aumentar la actividad de las enzimas del borde en cepillo como la sacarosa- isomaltasa (+82%), lactasa (+77%) y maltasa-glucoamilasa (+75%) -Buts y col., 1986-.
La dieta también puede estimular el crecimiento de ciertas bacterias como Bifidobacterium y Lactobacillus, lo que ayuda a aumentar la actividad enzimática de las proteasas, tripsina y lipasas -Hooper, 2001; Palmer y Rolls, 1983).
Finalmente, también ha sido demostrado que la microbiota intestinal favorece el desarrollo y la madurez de las vellosidades intestinales, aumentando la superficie de absorción, la producción de mucina y una digesta más ácida en comparación con las aves axénicas -Sakata y Setoyam, 1995-.
C. El papel de la microbiota a favor de la protección del animal
En un intestino sano, las bacterias residentes ocupan nichos de colonización intestinal.
La microbiota comensal suprime la proliferación de patógenos entéricos transitorios así como de bacterias oportunistas, a través de múltiples mecanismos -Kamada y col., 2014-:
Producción de metabolitos y competencia
La microbiota es capaz de producir metabolitos antimicrobianos como las β-defensinas o bacteriocinas que pueden alterar la permeabilidad de la membrana celular de las bacterias patógenas y conducirlas a la muerte o la incapacidad de desarrollarse -Brisbin y col., 2007-. Otro ejemplo es la capacidad de S.c. boulardii de producir proteasa -serina proteasa de 54 kDa- que inhibe las toxinas A y B de Clostridium difficile, o la fosfatasa -proteína fosfatasa de 63dKa), que ha mostrado ser eficaz contra el LPS de E. coli -Czerucka y col., 2002-.
Reducción del pH.
Las bacterias comensales tienen la capacidad de producir metabolitos como AGCC o lactato que disminuyen el pH intestinal. Estos ácidos también tendrán un efecto bacteriostático, ya que las bacterias patógenas requieren un pH intestinal más alto (5 a 5-8) para desarrollarse por completo -Blajman y col., 2015-.
Producción de mucus
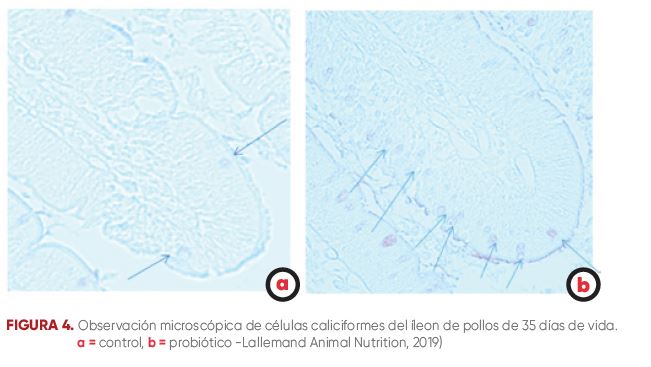
La superficie interna del intestino aviar está cubierta con una capa de mucus similar a un gel, que se forma a partir de la glucoproteína de mucina secretada por las células caliciformes. La capa de mucus evita que los microorganismos intestinales penetren en el epitelio intestinal y sirve como primera línea de defensa contra la infección -Johansson y col., 2013). Sakata y Setoyam (1995) demostraron que una microbiota intestinal saludable aumenta la producción de mucinas al proteger e influir en la producción de las glicoproteínas que forman el mucus (figura 4).
FIGURA 4. Observación microscópica de células caliciformes del íleon de pollos de 35 días de vida.
a = control, b = probiótico -Lallemand Animal Nutrition, 2019)
Función de barrera intestinal
La microbiota tiene un efecto sobre la protección de la barrera epitelial intestinal, estimulando a las proteínas intestinales específicas sprr2a que contribuyen al fortalecimiento del epitelio y la proliferación de células epiteliales -Dahan y col., 2003; Mumy y col., 2007; Hormann y col., 2014-.
Además, Czerucka y col. -2000- demostraron que S.c. boulardii refuerza las uniones estrechas epiteliales al reducir la capacidad de adherencia de las bacterias a las células epiteliales. Este efecto mejora la barrera epitelial intestinal y reduce la translocación bacteriana a los ganglios mesentéricos y a la sangre.
FIGURA 5.
D. La microbiota beneficia al sistema inmunitario
Las bacterias comensales también previenen la colonización y la infección de patógenos de manera indirecta al mejorar los mecanismos de defensa del huésped y la respuesta inmunitaria -Nunez y col., 2014-..
La investigación en animales criados en ausencia de microorganismos vivos conocidos como libres de gérmenes -GF-, reveló que la microbiota desempeña un papel crítico en el desarrollo de los órganos linfoides secundarios
Las bacterias comensales también previenen la colonización.
Este efecto es particularmente evidente en el tracto gastrointestinal donde se mencionan un tamaño menor de placas de Peyer y un número reducido de linfocitos T CD4+ y células plasmáticas productoras de IgA -Bauer y col., 1963; Hamada y col., 2002; Macpherson y col., 2001 Mazmanian y col., 2005; Smith y col., 2007; Talham y col., 1999-.
La microbiota juega un papel crucial en el desarrollo de la inmunidad del huésped
Los pollos libres de gérmenes contienen inmunoglobulina G -IgG- sérica inferior en comparación con los pollos criados convencionalmente -Thorbecke y col., 1957; Lecuyer y col., 2014-. La investigación mostró un desarrollo simultáneo del sistema inmunitario y de la microbiota intestinales.
Se sospecha que la microbiota intestinal desempeña un papel importante en el mantenimiento de la homeostasis inmunológica al prevenir la inflamación -Schaffner y col., 1974; Lei y col., 2015-. También se sabe que S.c. boulardii modula la inmunidad y la inflamación.
E. La levadura probiótica específica LEVUCELL SB
S.c. boulardii es una cepa de levadura que ha sido estudiada durante más de 60 años en salud humana y animal; y su modo de acción está altamente documentado por los científicos.
Es especialmente reconocida en salud humana por su eficacia contra trastornos gastrointestinales agudos y crónicos. Lallemand seleccionó su propia cepa de S.c. boulardii para aplicación animal y la registró bajo el nombre de CNCM I-1079 en el Instituto Pasteur -París, Francia-.
Caetano y col. -1986)- y Rodrigues y col. -2000- descubrieron que esta levadura viva activa la migración de monocitos y granulocitos hacia los ganglios linfoides. Además, mostraron que esta levadura aumenta los niveles de IgA en un 56,9% en el intestino delgado.
Varios estudios experimentales han mostrado que S.c. boulardii interfiere con las vías de señalización de la célula huésped y disminuye la expresión de citoquinas asociadas a la inflamación, como la interleucina 8 -IL-8), IL-6, IL-1β, el factor de necrosis tumoral alfa (TNF-α) y el interferón gamma (IFN- γ) – Dahan y col., 2003; Dalmasso y col., 2006b; Mumy y col., 2007 -. Estos estudios demostraron que esta levadura específica puede reducir la inflamación al bloquear el factor nuclear kappa B (NF-κB). El modo de acción de S.c. boulardii CNCM I-1079 está ampliamente documentado y ha sido estudiado en muchas especies monogástricas.
El probiótico ayuda a los pollos a hacer frente a situaciones difíciles al mejorar la morfología intestinal y estimular la actividad de los órganos inmunitarios.
EQUILIBRIO DE LA MICROBIOTA
S. c. boulardii CNCM I-1079 ayuda a al desarrollo de las bacterias benéficas y limita el crecimiento de ciertas bacterias indeseables en el intestino, en particular gracias a su capacidad para consumir oxígeno residual en el intestino (captación de oxígeno), lo que induce un entorno favorable para la flora benéfica. Esta levadura es capaz de producir proteasas y fosfatasas que pueden desactivar las toxinas provenientes de E. Coli, Clostridium y otros patógenos.
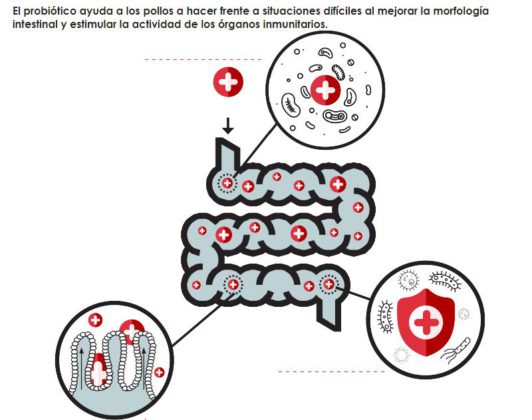
INTEGRIDAD INTESTINAL
S. c. boulardii CNCM I-1079 ayuda a mejorar la superficie de absorción intestinal aumentando la longitud de las microvellosidades y el ratio altura de vellosidades/profundidad de criptas. También ejerce un efecto positivo sobre la permeabilidad intestinal al reforzar las uniones estrechas entre las células epiteliales intestinales y reducir la capacidad de adherencia de las bacterias a las células epiteliales.
DEFENSAS NATURALES
Las defensas naturales de las aves se refuerzan con S. c. boulardii CNCM I-1079, a través de la modulación del sistema inmunitario local. Esto se traduce en la reducción de citocinas proinflamatorias como TNF-α, Il-1β, Il-6, Il-8, mientras que se mejora la producción de anticuerpos IgA protectores y citocinas antiinflamatorias (Il-10). Esta triple acción fortalece las funciones de barrera del intestino y mejora el equilibrio de la microflora, que da una mejor resistencia a los desafíos digestivos y un mejor rendimiento zootécnico
CONCLUSIONES
- El tracto digestivo juega un papel fundamental en la absorción de nutrientes y la protección contra los patógenos que podrían entrar en el cuerpo.
- Sus poblaciones microbianas juegan un papel importante tanto en los procesos digestivos como en el mantenimiento de la salud animal.
- La inclusión de un probiótico como S.c. boulardii para garantizar el equilibrio del ecosistema, así como una buena salud intestinal, es clave para lograr el mejor rendimiento y bienestar de las aves.
Fuente: Revista SELECCIONES AVÍCOLAS N#730 EQUIPO TÉCNICO DE LALLEMAND ANIMAL NUTRITION
Con el apoyo de:

Categorías
¿Quién es responsable en un caso de Toxiinfección alimentaria?
Leer
Clasificados
Leer
La “Mediterranean Poultry Summit 2020”, en Cordoba
LeerEl levante español acogerá las Jornadas Profesionales de Avicultura 2019
Leer
Legislación
Leer
In memoriam: Enrique García Martín
Leer
 PDF
PDF