Revistas
Revistas Archivo
Archivo









































Este artículo es parte de la edición de febrero, 2014
COCCIDIOSIS: LA ENFERMEDAD, CONSECUENCIAS Y TRATAMIENTO
E. del Cacho Malo (1) y Marc Pagès Bosch (2)
(1) Facultad de Veterinaria de Zaragoza
(2) Hipra Scientific, S.L.U.,Gerona

Fig. 1. Algunas de las características que definen a las especies de que infectan al pollo. (Adaptado de la tabla de Long y Reid, 1982).
La enfermedad
La coccidiosis aviar es una enfermedad parasitaria causada por protozoos del Phylum Apicomplexa, familia Eimeriidae. Aunque afecta a diversas especies de aves, es en el pollo de carne y gallina ponedora o reproductora donde alcanza la mayor repercusión económica. Es una enfermedad parasitaria que se produce mediante la ingestión de ooquistes esporulados, que dan lugar a un proceso de carácter clínico o subclínico, caracterizado por diarrea y descenso de las producciones.
El Phylum Apicomplexa incluye más de 4000 especies de protozoos alveolados que son todos parásitos obligados. Se caracterizan por tener un complejo apical formado por orgánulos especializados en el movimiento y la invasión de células del hospedador. La sistemática y taxonomía de los Apicomplexa está sometida a constantes cambios debido a la aparición de nuevas técnicas para el estudio del material genético, que se utilizan para construir nuevas relaciones filogenéticas. Los resultados obtenidos con estas técnicas ponen en duda las filogenias establecidas con datos basados en la morfología y características del desarrollo que definían en el pasado los grupos de especies.
En el pollo, Gallus gallus, se han citado15 especies diferentes. La mayoría de estas especies fueron descritas a partir de características morfológicas de ooquistes y la mayor parte de ellas no han vuelto a ser citadas. Desde los trabajos de Long y Reid – 1982 – se utilizan características intrínsecas a la fisiología y la patología del parásito para su clasificación taxonómica: el tiempo de esporulación, la localización del parásito en la mucosa intestinal, la localización y descripción de las lesiones, el número de esquizogonias, el tamaño de los esquizontes, el número y tamaño de los merozoítos, la inmunización cruzada con otras especies, las diferencias en composición enzimática y la densidad de flotación del ADN. En base a estos criterios Long y Reid publicaron un resumen con todos los parámetros mencionados para facilitar el diagnóstico de la coccidiosis aviar -fig. 1– en el que se mencionan nueve especies.
De estas nueve especies dos de ellas son todavía nomina dubia. La Eimeria hagani fue incluida en la lista de especies de Eimeria que infectan a Gallus gallus como de dudosa validez ya que desde su descripción por Levine –1938- no se ha vuelto a observar otro aislado de similares características. Eimeria mivati también es de validez dudosa. Su naturaleza como especie se pone en duda a partir del trabajo de Shirley y col. –1983- en el que después de estudiar con detalle un aislado de este parásito concluyeron que se trataba de una mezcla de ooquistes de E. acervulina y E.mitis. Son finalmente siete las especies que actualmente son aceptadas como especies de Eimeria capaces de infectar a Gallus gallus: E. acervulina, E. brunetti, E. maxima, E. mitis, E. necatrix, E. praecox y E. tenella.
El ciclo biológico de los protozoos del género Eimeria es similar al de otros protozoos de ciclo coccidial. De acuerdo con las descripciones realizadas por numerosos autores se pueden distinguir claramente dos fases:
a. Fase endógena: tiene lugar dentro del hospedador, y comprende las fases en las que los protozoos del género Eimeria se multiplican en el interior de células. Comprende dos fases de reproducción endógena, una asexual –esquizogonia- y otra sexual –gametogonia- que dará lugar a la formación del zigoto -ooquiste inmaduro- y al inicio de la fase exógena.
Los ooquistes esporulados -forma de resistencia con capacidad infectiva- son ingeridos por el hospedador, en cuyo tracto digestivo se produce el desenquistamiento. En una primera fase el dióxido de carbono y la acción mecánica de la molleja, producida por su musculatura y algunos elementos duros como restos de pienso o pequeñas piedras, consiguen alterar la estructura y la permeabilidad de la pared del ooquiste, lo que permite la acción de las sales biliares y la tripsina liberando los esporocistos.
En la segunda fase los esporozoítos abandonan los esporocistos mediante su propio movimiento estimulado por la acción de los ácidos biliares y las enzimas intestinales. Los ácidos biliares facilitan la entrada de enzimas intestinales en el ooquiste que ayudarán a activar el cuerpo de Stieda. Al mismo tiempo, los ácidos biliares estimulan la movilidad de los esporozoítos en el interior de los esporocistos y los abandonan a través del orificio que se produce como consecuencia de la alteración del cuerpo de Stieda.
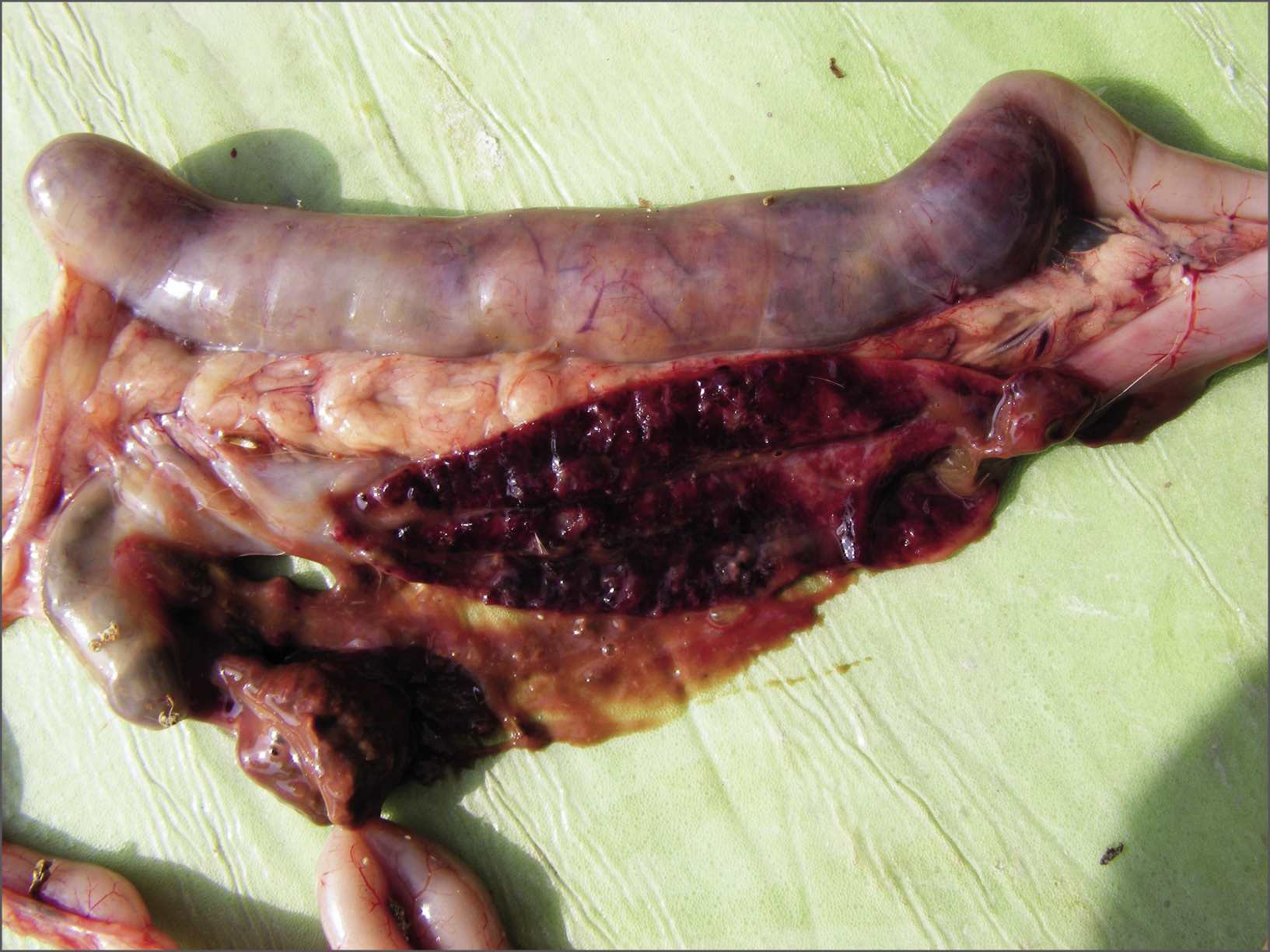
Lesiones de E. tenella en los ciegos. (Cortesía de HIPRA)
La edad de las aves tiene una influencia directa en el desenquistamiento de los ooquistes, cuya tasa es menor en el primer día de vida que en el tercero. Según Doran y Farr –1965-, la causa de esto podría ser la incapacidad de la parte muscular de la molleja para romper el ooquistes y la escasa cantidad de enzima pancreática presente en el tracto digestivo de los animales más jóvenes. Durante años se pensó que no era posible utilizar vacunas de parásito vivo -compuestas por ooquistes esporulados infectivos- en animales de entre uno y dos días de vida. Sin embargo, Williams y col. –1999- confirmaron que los pollitos de un día de vida son perfectamente capaces de desenquistar los esporozoítos, si bien es cierto que en animales jóvenes un 10% de los ooquistes pasa a través del intestino y son expulsados con las heces sin experimentar ninguna alteración en su pared.
Una vez que el esporozoíto ha conseguido penetrar en las célula epitelial forma la Vacuola Parasitofora y sufre un proceso de redondeamiento, transformándose en trofozoíto. Por divisiones nucleares múltiples -proceso de esquizogonia- el trofozoíto da lugar a los esquizontes de primera generación. Al final de la maduración del esquizonte se produce la rotura de la membrana de la célula hospedadora, permitiendo la liberación de los merozoítos que se observan en la luz de las criptas entre las 60 y 72 horas post-infección. Los merozoítos son formas muy similares a los esporozoítos en cuanto a estructura y capacidad infectiva. Son fusiformes y tienen un complejo apical que les permite desplazarse e infectar células epiteliales del intestino, dando lugar a diferentes generaciones de esquizontes -reproducciones asexuales-, que a su vez darán lugar a diferentes oleadas de merozoítos. El número de fases de reproducción asexual en las que se producen esquizontes que dan lugar a merozoítos –esquizogonias- es característico de cada especie y cepa de Eimeria -McDougald y Jeffers, 1976-. También tiene una estrecha relación con la patogenicidad y por lo tanto con la atenuación de las cepas de Eimeria. La última generación de merozoítos penetra en las células epiteliales de las criptas para desarrollar la gametogonia.
Los gamontes que se formarán a partir de los merozoítos, pueden ser de dos tipos:
- Macrogamontes -gametos femeninos-, que maduran para formar un único gameto, el macrogameto. La maduración no implica ninguna división, el macrogameto como el merozoíto es haploide y no lleva a cabo ninguna mitosis a diferencia de los esquizontes o microgamontes. En los macrogametos maduros destaca la presencia de los cuerpos formadores de la pared del ooquiste (BFWI y BFWII).
- Microgamontes -gametos masculinos-, sufren múltiples divisiones mitóticas para dar lugar a múltiples microgametos. Cuando maduran se convierten en bolsas que contienen en su interior gran cantidad de microgametos en distintas fases de desarrollo hasta completar su maduración -con dos flagelos y un núcleo haploide-.
La ruptura de la célula hospedadora permite la liberación de los microgametos, que abandonan la célula hospedadora y se desplazan hasta las células parasitadas con macrogametos. En estas células llevan a cabo la fertilización de los gametos femeninos y dan lugar a la formación del zigoto –ooquiste-. Las vesículas formadoras de la pared -BFWI y BFWII- migran a la superficie del zigoto, donde se fusionan para dar lugar a la pared del ooquistes. La pared final del ooquiste es una bicapa electrodensa formada por una capa externa, predominantemente lipídica, de 10 nm y por una capa glucoproteica en el interior de 90 nm. Esta bicapa es lo que confiere una resistencia extraordinaria al ooquiste inmaduro y maduro.
b. Fase exógena: En esta fase del ciclo se lleva a cabo una multiplicación asexuada por esporogonia, que tiene lugar fuera del hospedador. Brevemente, el proceso que se produce es el paso de un ooquiste diploide no esporulado y por tanto no infectivo a un ooquiste esporulado con ocho esporozoítos haploides infectivos.

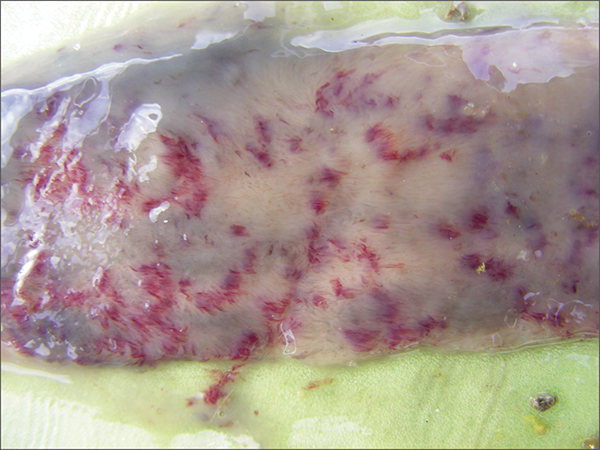
Infestación del duodeno por E. praecox. (Cortesía de HIPRA)
Los ooquistes son estadíos de resistencia del parásito y son eliminados sin esporular a través del flujo intestinal junto con las heces. El aspecto que ofrecen es el de una masa citoplásmica indiferenciada -esporonte- rodeada por una doble membrana. El proceso de esporogonia comienza cuando el ooquiste encuentra las condiciones adecuadas de oxigenación, temperatura y humedad. El esporoblasto inicial se divide primero en dos y posteriormente en cuatro, dando lugar a 4 esporoblastos que pasan de una forma triangular a redondeada, y se recubren de una doble membrana para formar los esporocistos. Estos esporocistos inmaduros sufren una división nuclear, para terminar el proceso de esporulación, con la formación en uno de los extremos del cuerpo de Stieda. De cada esporocisto se originan, dos esporozoítos o células infectantes. La esporulación de los ooquistes da como resultado ocho esporozoítos haploides a partir de un único ooquiste inicial diploide mediante un proceso de meiosis.
El periodo de esporulación está en función de la temperatura:
- 18 horas a 29ºC
- de 21 horas a 26-28ºC
- de 24 horas a 20-24ºC
- a temperatura ambiente son necesarias de 24-48 horas para completar la esporulación.
Long y Reid –1982- introducen el tiempo mínimo de esporulación como una de las características que definen a una especie de Eimeria. El problema es que factores como la humedad de la cama o la oxigenación son limitantes en el proceso de esporulación y por tanto en las explotaciones avícolas este parámetro presenta una gran variabilidad.
Las consecuencias
La afección intestinal inherente a toda infección por cualquiera de las especies de Eimeria citadas sigue un curso más o menos grave, que estará determinado por un gran número de variables como: la edad, el estado sanitario e inmunitario de las aves en el momento de la infección, el número de ooquistes ingeridos, las especies de Eimeria implicadas, etc. Distintos aislados de una misma especie han mostrado diferente patogenicidad intrínseca. En primoinfecciones realizadas con una misma cepa, su patogenicidad está directamente relacionada con la cantidad de ooquistes que ingiere el ave. Esta relación es limitada debido a la aparición de lo que se ha llamado el efecto multitudinario –“crowding factor”-, de manera que por encima de un nivel de infección las dosis mayores de ooquistes no producen mayores lesiones ni más cantidad de ooquistes.
La patogenicidad intrínseca de las especies de Eimeria parece estar directamente relacionada con el lugar de desarrollo, de manera que las especies más patógenas son las que penetran más profundamente en la mucosa intestinal. El poder patógeno de cada especie radica, principalmente, en las fases de esquizogonia y está en función de:
Factores dependientes del hospedador, como pueden ser la edad, las características genéticas, el estado de nutrición y, sobre todo, el estado de protección frente al parásito.
Factores ligados al propio parásito, como pueden ser el número de esquizogonias y el tamaño o la localización de los esquizontes. De esta forma se puede afirmar que E. tenella es la más patógena, seguida por E. necatrix y E. maxima.
La destrucción de las células epiteliales es el principal mecanismo de patogenicidad que subyace en la pérdida de la productividad de los animales infectados. La infección masiva determina la destrucción de un gran número de células epiteliales intestinales e incluso la destrucción de las vellosidades. Como consecuencia, se desencadena un síndrome de malabsorción, definido por la falta de absorción de nutrientes como aminoácidos, vitaminas y carotenos, cuya disminución determina pérdida de peso, descenso de la puesta y alteraciones en la calidad de la carne y de los huevos.
Cinco de las siete especies de Eimeria, que se establecen en el pollo y la gallina pueden considerarse como responsables de los cuadros más graves de coccidiosis clínica. Estas especies son:
- E. brunetti y E. maxima, que producen una diarrea sanguinolenta, o al menos con estrías sanguinolentas, que puede llegar a ser hemorrágica. En el caso de infecciones por E. maxima se observa una yeyunitis catarral que únicamente en procesos graves evoluciona a hemorrágica. En la mucosa del yeyuno se observan petequias que se producen como consecuencia del desarrollo de los ooquistes subepiteliales. El contenido es acuoso, de color rosáceo y puede mostrar coágulos de sangre. La E. brunetti produce una ileitis catarral que en procesos graves evoluciona a hemorrágica. En la mucosa del íleon y del recto se observan petequias que corresponden a la ruptura de los esquizontes subepiteliales.
- E. acervulina se asocia con enteritis catarral y produce, principalmente, diarreas mucosas, que originan detrimento en la ganancia de peso. En los casos clínicos producidos por E. acervulina se observa una duodenitis catarral que puede evolucionar a mucosa con un contenido de color amarillento. En la mucosa del duodeno se observan lesiones puntiformes que pueden fusionarse hasta formar placas que se orientan en sentido transversal al intestino y con aspecto escaleriforme. Estas lesiones corresponden a acúmulos de macrogametos y ooquistes inmaduros. Tras la liberación de los ooquistes las vellosidades del duodeno se observan desprovistas de células epiteliales.
- E. tenella y E. necatrix, productoras de hemorragias a partir de finales del 4º y principios del 5º día post infección, que se asocian a la maduración de los esquizontes de segunda generación y causan una elevada tasa de mortalidad. Las lesiones producidas por E. necatrix se localizan en el yeyuno. Se observa yeyunitis catarral, que frecuentemente evoluciona a hemorrágica. En la mucosa del yeyuno se observan lesiones puntiformes y blanquecinas que corresponden a acúmulos de esquizontes subepiteliales que al romperse producen petequias e incluso hemorragias en procesos muy graves. En la necropsia de los animales que muestran diarrea hemorrágica como consecuencia de una infección por E. tenella se observa tiflitis hemorrágica con los ciegos notablemente dilatados. En la mucosa de los ciegos se observan desde petequias a hemorragias, dependiendo de la gravedad del proceso. En la luz de los ciegos se detecta un contenido hemorrágico frecuentemente con coágulos de sangre. Estas lesiones se producen como consecuencia de la ruptura de los esquizontes subepiteliales. Los animales que superan esta fase muestran en los ciegos un contenido solidificado que se definen como moldes cecales y que corresponden a la coagulación del contenido cecal que se produjo como consecuencia de la esquizogonia. Mediante la realización de cortes histológicos se puede observar que la mucosa cecal está completamente destruida y que los moldes cecales están constituidos por acúmulos de restos celulares entre los que destacan los glóbulos rojos. Los brotes clínicos producidos por E. tenella conllevan una alta tasa de mortalidad. Sin embargo, los animales que sobreviven eliminan en las heces los moldes cecales que son expulsados como estructuras alargadas y cilíndricas, de color amarillento claro. Puesto que la gravedad de las lesiones en una primoinfección está relacionada con el número de ooquistes ingeridos, se ha realizado la gradación de las lesiones en función de la presencia de petequias, hemorragias o moldes cecales – Johnson y Reid, 1970 -.
La mayoría de los brotes clínicos se deben a infecciones mixtas de varias especies de Eimeria y, por tanto, se observan lesiones en diferentes zonas del intestino. La interacción entre especies de coccidios puede desarrollarse siguiendo cursos distintos, así las especies que parasitan la misma región intestinal – E. praecox y E. acervulina – compiten y el efecto combinado no es mayor que cuando la infección la produce una sola especie, pero entre especies que parasitan distinta región intestinal -E. brunetti y E. acervulina- el efecto patógeno de la combinación es mucho mayor.
Existen factores que no están directamente relacionados con la patogenicidad intrínseca de las especies de Eimeria, pero que pueden influir en el pronóstico de una infección. Entre estos factores cabe destacar:
- La edad de las aves: los animales mayores que han sido mantenidos libres de coccidios durante su crianza son más receptivos a la infección que los jóvenes.
- Diferencias genéticas observadas en la resistencia a la coccidiosis entre distintas razas o estirpes de aves. Se han descrito estirpes que parecen presentar una mayor resistencia a la coccidiosis, aunque todas las estirpes comerciales parecen presentar la misma receptividad.
- Interacciones con otras patologías: deficiencias nutricionales, bacterias, virus, parásitos y micotoxinas. Estas interacciones son difíciles de demostrar en el campo, pero crece la evidencia indirecta de que son cada vez más importantes a medida que los avances en genética y nutrición producen aves de crecimiento más rápido.
Foco de lesiones de E. maxima. (Cortesía de HIPRA)
El tratamiento
Una gran cantidad de fármacos manifiestan la capacidad de dificultar la multiplicación de Eimeria, principalmente inhibiendo la esquizogonia y, por tanto, las manifestaciones clínicas de la enfermedad. Con la finalidad de regular la utilización de los compuestos medicinales usados en la industria aviar la UE prohibió una larga lista de productos con efecto coccidiostático. El Reglamento (CE) Nº 2205/2001 limita la lista de coccidiostáticos autorizados como aditivos a las siguientes:
- Robenidina
- Halofuginona
- Diclazuril
- Decoquinato
- Narasina/Nicarbacina
- Lasalocid
- Salinomicina sódica
- Maduramicina de amonio
- Monensina de sodio
- Narasina
Posteriormente se ha permitido la utilización de la nicarbacina como aditivo –REGLAMENTO (UE) Nº 875/2010- y la utilización de Clinacox, Maxiban y Monteban sin periodo de retirada -REGLAMENTO (CE) Nº 976/2008, REGLAMENTO (UE) Nº 885/2010 y REGLAMENTO (UE) Nº 884/2010-. La vía de administración más utilizada es añadir los fármacos a la alimentación de las aves a la concentración establecida para cada compuesto activo.
Las especies de Eimeria responsables de la coccidiosis son capaces de desarrollar resistencias frente a todos los fármacos descritos. La formación de cepas resistentes se ve facilitada cuando se utilizan dosis bajas durante largos períodos de tiempo. Cuando se producen brotes clínicos debido al desarrollo de resistencias es recomendable realizar una prueba de sensibilidad anticoccidiósica para establecer el tratamiento adecuado, aunque en la mayoría de las veces es suficiente con cambiar el coccidiostático. La prueba de sensibilidad anticoccidiósica se lleva a cabo mediante la administración experimental de las cepas sospechosas de presentar un cierto grado de resistencia. Posteriormente, las aves se someten a tratamiento con diferentes coccidiostáticos y se valora la eficacia de cada uno de ellos mediante la cuantificación de los parámetros parasitológicos -lesiones intestinales, mortalidad y excreción de ooquistes- y la cuantificación de los parámetros productivos -ganancia en peso, índice de conversión, etc-.
Todos los fármacos descritos se pueden utilizar en un tratamiento preventivo o curativo. En el tratamiento preventivo, los fármacos se añaden al pienso de la alimentación de las aves con la finalidad de prevenir la multiplicación de las especies de Eimeria responsable de la coccidiosis aviar. Para evitar la aparición de resistencias se han propuesto dos pautas de tratamiento preventivo, que son:
- El sistema rotacional, que propone cambiar el coccidiostático cada 4-6 meses de producción.
- El sistema dual, que consiste en cambiar el coccidiostático a la mitad de un ciclo de cría, entre 21 y 25 días de vida.
En el curso de los tratamientos preventivos, si no hay resistencia a los coccidiostáticos, las aves no desarrollan inmunidad y, por tanto, cuando dejan de recibir coccidiostáticos presentan una alta receptividad a la infección. Si se genera inmunidad frente Eimeria en el curso de la quimioprofilaxis es debido a que existen poblaciones de Eimeria resistentes al coccidiostático que dan lugar a una tasa antigénica suficiente para generar una respuesta inmune eficaz frente al parasito.
En el tratamiento curativo se aplica en el caso de que se desencadene un brote clínico de coccidiosis aviar. Se pueden utilizar los mismos fármacos que se emplean como aditivos o algunos otros considerados como medicamentos. Generalmente se añaden al agua de bebida de las aves durante 3-5 días. Se administran dosis que varían desde el 0,1 % – sulfamidas -hasta el 0,01 % -Amprolio-.
El principal problema de la quimioprofilaxis es que si se mantiene la relación del parasito con el coccidiostático es cuestión de tiempo que aparezcan poblaciones resistentes al fármaco utilizado. Pero, la resistencia a coccidiostáticos no es genéticamente estable, de forma que cuando se deja de utilizar el producto las poblaciones resistentes desaparecen y aumentan las poblaciones sensibles. Por eso algunos autores -Chapman y col. 2010- han propuesto que la mejor estrategia frente a la coccidiosis es la alternancia entre coccidiostáticos y vacuna.
Las vacunas utilizadas en Europa están constituidas por el parásito vivo atenuado y las características de patogenicidad e inmunogenicidad de las cepas vacunales vienen determinadas en European Pharmacopoeia –2013-. Las cepas vacunales se atenúan reduciendo el periodo de prepotencia mediante una disminución del tamaño y número de esquizontes, lo que implica menor patogenicidad y menor capacidad de multiplicación. Las vacunas constituidas por el parasito vivo atenuado presentan un excelente efecto de inmunización frente a Eimeria pero presentan varios problemas, siendo quizás el más importante el que la inmunidad desencadenada por ellas es específica para las cepas vacunales. Eso quiere decir que las vacunas no inmunizan frente a las especies de Eimeria que no están incluidas en la formula vacunal. También se ha descrito que las cepas de campo pueden presentar una gran diversidad antigénica con respecto a las cepas vacunales y por tanto desencadenar una coccidiosis subclínica en animales vacunados.
Con la finalidad de solucionar los problemas planteados por las vacunas constituidas por parásito vivo se están desarrollando vacunas frente a la coccidiosis aviar mediante el aislamiento de antígenos, la inmunización con plásmidos, constituidos con fragmentos de ADN del parásito y la utilización de exosomas. A juzgar por los resultados expuestos en las publicaciones científicas generadas a partir de estas vacunas experimentales la utilización de exosomas parece la estrategia más esperanzadora. Estas vacunas están constituidas por vesículas secretadas por las Células Dendríticas que se pueden recubrir con antígenos de diversos patógenos para desarrollar una vacuna polivalente capaz de inmunizar frente a virus, bacterias y todas las especies de Eimeria – del Cacho y col., 2011; del Cacho y col. 2012-.
Referencias bibliográficas
(Se enviarán a quienes las soliciten)
Con el apoyo de:

Categorías

S.P. Veterinaria
Leer Tashia
Leer Ingenieria Avicola
Leer Camagán
Leer Chore-Time
Leer Big Dutchman
Leer
 PDF
PDF